3.04 – Ciclo di Krebs
Ultimo aggiornamento: 30/07/2023
Il ciclo di Krebs deve il suo nome ad una sua importante caratteristica: ha un andamento circolare. Infatti, il primo composto che reagisce con l’acetil-CoA entrante, è l’ossalacetato che rappresenta anche il prodotto finale del ciclo. Nel mezzo, si susseguono una serie di otto reazioni da cui si passa da un composto a sei atomi di carbonio ad uno a cinque e infine di nuovo uno a quattro atomi di C (ossalacetato).
Durante quattro di queste otto tappe avvengono delle ossidazioni con formazione dei coenzimi ridotti NADH e FADH2 (3 NADH e 1 FADH2 per ogni molecola di piruvato). Si formano inoltre anche due molecole di CO2 e una di GTP (molecola simile all’ATP).
Nei procarioti, glicolisi e ciclo di Krebs avvengono nello stesso compartimento: il citoplasma. Negli eucarioti invece assistiamo ad un cambio location. Il piruvato passa prima attraverso i pori sulla superficie della membrana mitocondriale esterna e poi diffonde nella matrice grazie al trasportatore del piruvato mitocondriale (MPC). Qui viene ossidato ad acetil-CoA grazie ad un complesso multienzimatico che prende il nome di complesso della piruvato deidrogenasi (PDH).
Durante la reazione catalizzata dall’enzima, un gruppo carbossilico viene rimosso sotto forma di anidride carbonica (CO2) e l’elettrone sottratto viene immagazzinato dal NAD. Infine, il gruppo acetilico formato si unisce al coenzima A.
TAPPA 1
Nella prima tappa effettiva del ciclo di Krebs, l’acetil-CoA va incontro ad idrolisi e il gruppo acetilico si lega al gruppo carbonilico dell’ossalacetato. La reazione è catalizzata dalla citrato sintasi, che lega prima l’ossalacetato, responsabile di una modificazione conformazionale dell’enzima, e poi il gruppo acetil-CoA. Nel sito attivo si forma dapprima un composto intermedio, il citroil-CoA, dopodiché una seconda modificazione conformazionale porta all’idrolisi del coenzima A e alla formazione del citrato.
TAPPA 2
Il citrato viene trasfromato in isocitrato mediante una reazione che porta ad un composto intermedio (cis-aconitato). Lo scopo della reazione è spostare il gruppo -OH dal carbonio in posizione 3 al carbonio in posizione 4 attraverso l’eliminazione e poi l’aggiunta di una molecola d’acqua. L’enzima che catalizza la reazione prende il nome di aconitasi.
TAPPA 3
A questo punto si passa da un composto a sei atomi di carbonio ad un composto che ne ha cinque: l’isocitrato deidrogenasi catalizza la rimozione di un gruppo carbossilico con formazione di α-chetoglutarato. La reazione avviene in più step. Inizialmente il gruppo -OH posto precedentemente sul carbonio 4 viene ossidato. Successivamente l’intermedio viene decarbossilato a livello del carbonio 3 ed infine, il composto enolico che si forma viene riarrangiato per formare α-chetoglutarato.
TAPPA 4
L’ ⍺-chetoglutarato è convertito in succinil-CoA dall’enzima complesso dell’⍺-chetoglutarato deidrogenasi con formazione di NADH+H+.
Questa reazione è simile a quella dell’acetil-CoA. L’enzima è un complesso enzimatico come la piruvato deidrogenasi e in entrambe le reazioni si forma NADH+H+.
Da questa reazione viene prodotta anche un’altra molecola di CO2. Fino a qui sono state prodotte due molecole di CO2 e due di NADH+H+.

Reazione 5
Il succinil-CoA è convertito in succinato dall’enzima succinil-CoAsintetasi. Il succinil-CoA è un tioestere ad alta energia, che verrà convertito in un succinilfosfato. Il fosfato verrà poi donato all’GDP per produrre GTP e succinato. Dal GTP si formerà poi una molecola di ATP.

Reazione 6
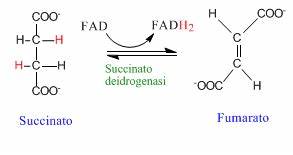
Il succinato è convertito dall’enzima succinato deidrogenasi in fumarato. Con questa reazione inizia la fase di riconversione ad ossalacetato attraverso varie tappe che porteranno all’introduzione finale di un carbonio carbonilico.
Per fare ciò con la reazione di deidrogenazione otteniamo innanzitutto il doppio legame, che verrà poi idratato ed ossidato così da ottenere il gruppo chetonico dell’ossalacetato.
Pertanto le prossime tre reazioni avranno come fine ultimo quello di produrre ossalacetato oltre che ovviamente ATP. La Particolarità di questa reazione è che determina la riduzione del FAD a FADH2 e non del NAD, perché l’energia associata alla reazione non permette la riduzione del NAD. È questa infatti l’unica reazione del ciclo di Krebs in cui l’enzima utilizza il FAD, al quale è legato covalentemente mediante un residuo di istidina.
L’enzima succinato deidrogenasi è importante perché è coinvolto anche nella catena di trasporto degli elettroni, pertanto è annidato nella membrana mitocondriale costituendo il complesso II di tale catena. Gli elettroni caricati sul FADH2 vengono quindi immessi direttamente nella catena di trasporto degli elettroni, grazie al legame stabile tra l’enzima e il cofattore stesso. Fin qui abbiamo prodotto, 2 CO2, 2 NADH+H+, una molecola di ATP dal GTP e un FADH2.
Reazione 7
Il fumarato è convertito in malato dall’enzima fumarasi. È una reazione di idratazione in trans del doppio legame che porta alla formazione di un gruppo alcolico.
L’enzima è in grado di aggiungere il gruppo OH solo da un lato pertanto otteniamo solo l’ L-malato.

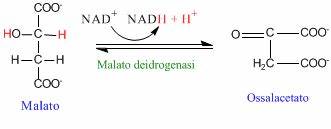
Reazione 8
Il malato è convertito in ossalacetato dall’enzima malato deidrogenasi. È una reazione di ossidazione del gruppo OH che comporta la formazione del gruppo chetonico.
L’enzima utilizza come cofattore il NAD+, pertanto si ha la produzione di un’altra molecola di NADH+H+.
Con quest’ultima reazione si è ottenuto nuovamente l’ossalacetato pertanto il ciclo può ricominciare.
Regolazione del ciclo di Krebs
Il ciclo di Krebs è regolato su tre tappe esoergoniche:
1. Reazione di formazione del citrato catalizzata dalla citrato deidrogenasi, l’enzima viene inibito dal NADH, anche se meno rispetto all’isocitrato deidrogenasi, e dal suo prodotto, mentre è attivato dalle alte concentrazioni di ossalacetato.
2. Reazione di formazione dell’ ⍺-chetoglutarato catalizzata dall’isocitrato deidrogenasi, l’enzima è inibito dal NADH e dal suo prodotto, oltre che dalle alte concentrazioni di ATP.
3. Reazione di formazione del succinil-CoA catalizzata dall’ ⍺-chetoglutarato deidrogenasi, l’enzima è inibito dal NADH e dal suo prodotto. Inoltre il succinil-CoA compete con l’acetil-CoA andando a inibire a distanza la citrato deidrogenasi (inibizione retroattiva competitiva «a feedback»)

Metabolismi che utilizzano gli intermedi del ciclo di Krebs
• Ossalacetato: viene utilizzato per la gluconeogenesi, che è un processo inverso alla glicolisi. L’ossalacetato infatti può essereri convertito in fosfoenolpiruvato nel citoplasma. Per poter uscire dal mitocondrio si ha prima la conversione dell’ossalacetato in malato, che può attraversare la membrana mitocondriale. Una volta nel citoplasma viene riconvertito in ossalacetato e quest’ultimo in fosfoenolpiruvato. Da qui in avanti si svolgono tutte le reazioni inverse della glicolisi sino al glucosio.
• Acetil-CoA: viene utilizzato nella sintesi dei lipidi, mentre è prodotto dalla loro degradazione. Tuttavia mentre la degradazione avviene nella matrice del mitocondrio, la sintesi si svolge nel citoplasma pertanto si ha la necessità di trasportare l’acetil-CoA fuori dal mitocondrio. Per ottenere ciò si forma, dall’acetl-CoA e ossalacetato, il citrato che passa dal mitocondrio al citoplasma. Qui grazie all’enzima ATP-citrato-liasi, il citrato viene scisso nuovamente in ossalacetato e acetilCoA che può essere così utilizzato per la sintesi dei lipidi.
• Ossalacetato e ⍺-chetoglutarato: sono entrambi degli ⍺-chetoacidi con gruppi carbossilici alle estremità e un gruppo chetonico. Se prendiamo in considerazione due amminoacidi, acido aspartico e acido glutammico, sono entrambi definiti monoammino-dicarbossilici dato che hanno un gruppo amminico nella catena carboniosa e due gruppi carbossilici alle estremità. In seguito a deaminazione («perdita») dei loro gruppi amminici otteniamo rispettivamente ossalacetato e ⍺-chetoglutarato, con la reazione inversa possiamo invece riottenere nuovamente gli amminoacidi. Questa reazione di conversione dell’amminoacido nel suo chetoacido corrispondente, e viceversa, è detta transaminazione.
• Succinil-CoA: viene utilizzato nella sintesi dell’eme. Infatti essa parte dalla sintesi dell’acido 5- (oppure delta)-amminolevulinico (dALA) dall’amminoacido glicina e dal succinil-CoA.
Reazioni che forniscono gli intermedi al ciclo di Krebs
Per rifornire Ossalacetato: il piruvato reagisce con la CO2, in presenza di ATP e acqua, secondo la seguente reazione, catalizzata dall’enzima piruvato carbossilasi:
Piruvato + CO2 + ATP + H2O ossalacetato + ADP + Pi
Per rifornire succinil-CoA: si attiva la degradazione degli acidi grassi oppure di alcuni amminoacidi (isoleucina, metionia evalina).
Per rifornire Ossalacetato e ⍺-chetoglutarato: si ha la reazione di transaminazione.
BILANCIO DEL CICLO DI KREBS
in totale dal ciclo di Krebs si sono ottenute:
2 CO2, 3 NADH+H+
1 ATP
1 FADH2
il tutto moltiplicato per due dal momento che partiamo da due molecole di piruvato prodotte dalla glicolisi
quindi 4 CO2, 6 NADH+H+, 2 ATP e 2 FADH2
Altri 2 NADH+H+ sono stati invece prodotti nella reazione del piruvato ad acetil-CoA.
Da ogni molecola di cofattore prodotto si otterranno altre molecole di ATP (2,5 per ogni NADH e 1,5 per ogni FADH2) che si andranno ad aggiungere al bilancio energetico finale.
Esercizio 1
Il punto di unione del ciclo di Krebs e della glicolisi è rappresentato da
A acido piruvico
B acido ossalacetico
C glucosio
D acetil-CoA
E nessuna delle risposte è corretta
Correzione commentata
risposta corretta lettera D: Il punto di unione del ciclo di Krebs (o ciclo degli acidi tricarbossilici) e della glicolisi è rappresentato dall’acido piruvico. L’acido piruvico è il prodotto finale della glicolisi, che è la prima tappa del metabolismo del glucosio. Durante la glicolisi, il glucosio viene convertito in due molecole di acido piruvico. Successivamente, l’acido piruvico entra nella matrice mitocondriale, dove viene trasformato in acetil-CoA, una molecola composta da due atomi di carbonio. L’acetil-CoA è il substrato che avvia il ciclo di Krebs. Durante il ciclo di Krebs, l’acetil-CoA viene ulteriormente ossidato e degradato, producendo CO2, energia sotto forma di ATP e molecole di ridotto come il NADH e il FADH2.
Quindi, l’acido piruvico è il punto di unione tra la glicolisi e il ciclo di Krebs, poiché l’acido piruvico prodotto durante la glicolisi viene convertito in acetil-CoA per entrare nel ciclo di Krebs.
Esercizio 2
Ogni ciclo di Krebs quanti gruppo acetile consuma ?
a. uno
b. due
c. tre
d. quattro
e. zero
Correzione commentata
risposta corretta A: Durante il ciclo di Krebs, l’acetil-CoA, che è il substrato iniziale, viene completamente ossidato e degradato, producendo energia sotto forma di ATP, molecole di ridotto come il NADH e il FADH2, e liberando anidride carbonica (CO2) come prodotto di scarto. L’acetil-CoA fornisce un gruppo acetile, composto da due atomi di carbonio, che viene completamente ossidato nel corso del ciclo di Krebs. Quindi, in ogni ciclo di Krebs, viene consumato un gruppo acetile, che corrisponde a due atomi di carbonio. La risposta corretta è quindi l’opzione a) uno.